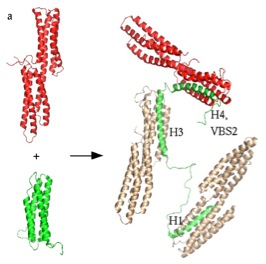
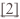Talin –vinculin helix interactions are dominated by hydrophobic side chains
Talin domain (green) unfolds and individual helices interact with vinculin binding sites


![A new area of interest for our group is the development of proteomimetics, stable artificial synthetic structures that will mimic protein secondary and tertiary structures.
Controlling protein-protein interactions (PPIs) represents a significant contemporary challenge since protein surfaces are often large and solvent exposed with non-contiguous binding regions. Rationally designed ‘proteomimetic’ molecules have shown promise for successfully mimicking secondary protein structures by projecting key hydrophobic residues for binding at ’hotspots’ identified from structural and mutation studies.
We are developing new approaches to the rational design of proteomimetics based on a detailed knowledge of structures protein-protein complexes obtained in the Liverpool NMR Centre for Structural Biology.
Talin and focal adhesion
Talin is a modular multidomain cytosolic protein that couples the integrin receptor to the cytoskeleton and thus plays a key role in cell adhesion. It is the central hub for focal adhesion complex formation bearing binding sites for a number of interaction partners including, integrins, RIAM, PIP2, actin and vinculin.[1]
Many of the known interactions involve helix-helix contacts between cognate protein ligands and the 4 and 5-helix bundle domains of talin. For examle, talin contains a number of vinculin binding sites (shown as red elipses above) that are released upon unfolding of the helical bundles. A number of the binding sites are shared by more than one binding partner and the specificity is not yet understood. We aim to construct stable proteomimetics that can selectively disrupt or stablise these interactions as probes to faciliate further detailed understanding about the role of talin and its complexes in cell adhesion.
Frequenin/Neuronal Calcium Sensor 1 (NCS-1) protein-protein interactions
Frequenin (Frq) or Neuronal Calcium Sensor 1 (NCS-1) is a member of a subclass of the calmodulin superfamily of proteins that has a range of binding partners.[2] Many of these proteins utilise a calcium-myristoyl switch and function as calcium sensors. Members of the family include recoverin and GCAP, which are involved in the control of visual signal transduction. In yeast, frequenin binds calcium and the resulting conformational change releases an N-myristoyl group which subsequently localises the frequenin to the membrane where it interacts with and activates phosphatidyl inositol 4-kinase (PiK1).[3] In mammalian cells NCS-1 is important for neuronal function and regulates the orthologous PiKIIIβ.[4] Frequenin/NCS-1 protein-protein interactions involve side-on interactions of several helices from the kinase with an open hydrophobic surface on frequenin, revealed following calcium binding.
We are interested in developing selective inhibitors against NCS-1 proteins in order to further understand the specificity and function of its interactions with various ligands.](Protein-Protein%20Interactions_files/shapeimage_3.png)